Klas Monoplacophora – die Oer-eenskulpiges
Die naam Monoplacophora, of monoplakofore, verwys na die enkel skulp wat hierdie groep het – in kontras met die talle skulpe van die Polyplacophora, of poliplakofore wat ook in die volksmond as kewerslakke bekend staan. Ten spyte van die ooreenkoms tussen hulle doppe met dié van klipmossels, is die monoplakofore en Gastropoda of buikpotiges slegs baie verlangs verwant. Vir lank het mense gedink dat die Polyplacophora, of kewerslakke, die mees primitiewe weekdiere is aangesien hulle ‘n paar primitiewe kenmerke soos series-gerangskikde spiere en natuurlik die doppe wat mens aan segmente laat dink. Dit was voor die wurmagtige weekdiere – die Aplacophora, of aplakofore, (die naam bedoel “geen skulp”) ontdek is, wat nog veel primitiewer blyk te wees.
Die rede vir hierdie soeke na primitiwiteit gaan oor die obskure herkoms van weekdiere. Weekdiere was reeds in die Kambriese Tydperk teenwoordig. Net soos die gesegmenteerde wurms en die geleedpotiges, is weekdiere ook euselomate, maar anders as die gesegmenteerde wurms, toon die weekdiere geen opsigtelike segmentasie nie, wat ‘n taksonomiese probleem is, aangesien alles daarop dui dat die seloom en segmentasie saam ontstaan het en dus dat die voorouers van die weekdiere en geleedpotiges van voorouers, wat naasverwant aan die gesegmenteerde wurms was, afgestam het.
Dit is verstaanbaar dat, soos wat organismes se behoeftes oor miljoene jare verander en hulle by veranderende omstandighede en lewenswyses moet aanpas, sekere kenmerke verdwyn of verander het. Een so ‘n kenmerk was dat die septa, wat as afskortings tussen die segmente in gesegmenteerde wurms voorkom, in weekdiere en geleedpotiges verdwyn het. Alhoewel geleedpotiges serie-gerangskikte eksoskeletsegmente, spiere en organe het, het hulle nie meer septa tussen hulle segmente nie. Meeste weekdiere het bykans alle tekens van serie-gerangskikte organe en strukture verloor, buiten soos genoem, die kewerslakke wat steeds serie-gerankskikde spiere en veelvoudige dekplate het.
Monoplakofore was reeds vanaf die 18de Eeu as fossiele bekend. Aangesien niemand lewende verteenwoordigers van hierdie groep voor 1952 ontdek het nie, was daar aanvaar dat hulle in die Devoon Periode omtrent 375 miljoen jaar gelede uitgesterf het. Daar was dus groot opgewondenheid toe die Deense navorsingskip Galathea lewende monoplakofore op die seevloer op ‘n diepte van 3 570m van die kus van Costa Rica ontdek het. Dit was as een van die grootste biologiese ontdekkings van die 20ste Eeu beskou – op dieselfde vlak van belangrikheid as die ontdekking van die selekant in 1938 en die Wollemi Den in Australië in 1995 – want talle vrae ten opsigte van die ontstaan en evolusie van weekdiere kon nou beantwoord word.
Reeds van die staanspoor was paleontoloë oortuig dat die fossielskulpe bewys dat die monoplakofore ‘n aparte en baie primitiewe groep weekdiere is en nie buikpotiges (Gastropoda) nie, ondanks die oppervlakkige ooreenkoms tussen die monoplakofore se skulpe en dié van die klipmossels. Monoplakofore se skulpe wissel tussen 3 en 37 mm lank. Die groot verskil was dat daar twee rye holtes aan die binnekant van monoplakofore se skulpe was. Hierdie holtes was die aanhegtingsplekke van die spiere van die voet. Gastropoda het een groot spier wat die voet opmaak, maar in die kewerslakke is daar ook sulke reekse voetspiere.
Die 35 bekende ekstante monoplakofoorspesies word in sewe genera en in twee families verdeel (Ponder et al. 2020). Ekstante monoplakofore kom vanaf 174 meter diep of dieper op die seevloer van die kontinentale bank, die kontinentale helling en in die abissale sone tot op dieptes van ongeveer 6 500 meter voor. Die oudste monoplakofore het in die Kambrium Periode hoofsaaklik in vlak oseane voorgekom, terwyl hul nasate meer algemeen in die sagte modder diep op die seevloer voorgekom het.
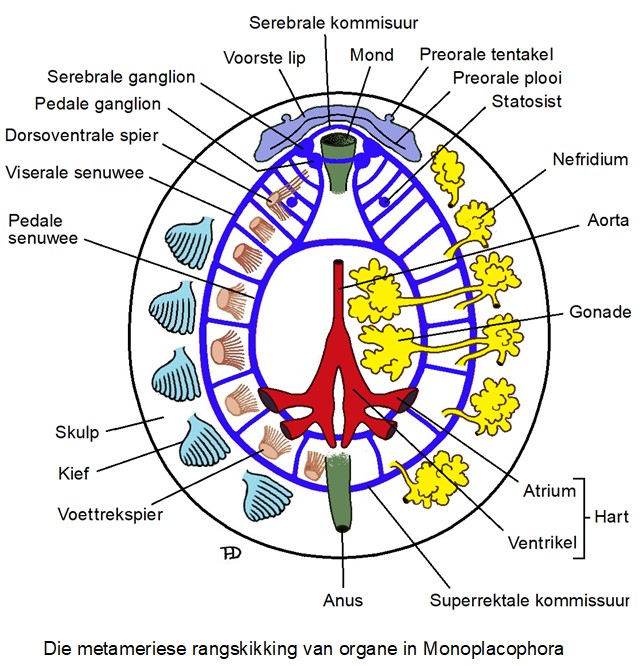
Na die ontdekking van die eerste ekstante monoplakofore kon die hipotese dat die reeks gate binne die dop vir spieraanhegtingsplekke gebruik was, bevestig word. Monoplakofore het ook verskeie ander organe wat in series gerangskik is. Hierdie verskynsel word in die afwesigheid van septa pseudometamerisme genoem (as daar segmente was wat van mekaar deur septa geskei was – soos wat mens in die gesegmenteerde wurms (Annelida) sien, sou mens dit metamerisme genoem het).
Hierdie herhaling van organe is nie uniek aan die monoplakofore nie en word ook in kewerslakke en protobrangiese tweekleppiges en die koppotige Nautilus aangetref. Nautilus het twee pare voetspiere, niere, kiewe en atria. In monoplakofore is daar nul tot twee pare aurikels, drie tot ses pare kiewe en drie tot sewe pare niere. Daar is agt skulpplate in kewerslakke en hul het verskeie pare voetspiere en kiewe.
Daar is twee hoofteorieë oor hierdie herhaling van organe of pseudometamerisme in weekdiere. Die eerste is dat dit ‘n primitiewe of plesiomorfiese, eienskap is wat van hul gesegmenteerde voorouers (soos Annelida – die gesegmenteerde wurms) oorgeërf is (Wingstrand, 1985). Die tweede is dat dit ‘n sekondêre ontwikkeling is wat nie slegs in weekdiere voorkom nie, maar in talle ander diertaksa, buiten gesegmenteerde wurms, soos die gonade van Nemertea, die proglottiede van Cestoda en die spiere van Briozoa en Brachiopoda (Haszprunar & Schaefer, 1996; Giribet 2016).
Monoplakofore verskil anatomies heeltemal van buikpotiges, maar kom in baie opsigte met kewerslakke ooreen. Monoplakofore het, soos die kewerslakke, agt paar series, of segmentaal-gerangskikte, dorso-ventrale spiere, wat ek hier voetspiere noem, wat die skulp aan die lyf vasanker. Die senuweestelsel van monoplakofore is soos dié van die kewerslakke, baie eenvoudig en daar is klein ganglions of ganglionsa in die kop, waarteenoor die ganglionsons van buikpotiges en veral koppotiges heelwat groter is en in laasgenoemde ‘n brein vorm.
Skulp
Die apeks (bopunt) van die skulpe van ekstante en uitgestorwe monoplakofore is vorentoe-gerig wat in sekere spesies selfs verder vorentoe as die onderste rand van die skulp uitsteek. Die vorm van die ronding van die skulp wissel in verskillende spesies van amper koeëlrond tot peervormig, terwyl die hoogte van die skulpe van platterig tot ‘n lae keëlvorm wissel. Ekstante spesies se skulpe is tipies minder as 1 cm lank, terwyl meeste fossielspesies se skulpe 2-5 cm lank is. Die buitenste oppervlakte van die dop is met ‘n dun periostrakum bedek wat met dié van ander weekdiere se skulpe eerder as dié van kewerslakke ooreenstem. ‘n Boonste prismatiese skulplaag en ‘n binneste gefolieërde laag is onder die periostrakum van meeste monoplakofore geleë. Die binneste laag van die skulp is met ‘n laag gefolieërde aragoniet uitgevoer.
Kop en mantel
Die kop van monoplakofore, soos dié van kewerslakke is klein en onopmerklik en slegs uitkenbaar weens die posisie van die mond wat ondertoe open. ‘n Transversale groef skei die kop van die voet. Daar is geen nek tussen die eenvoudige kop van die monoplakofore en die res van die liggaam nie. Die voorste deel van die kop bestaan uit die voorste lip en kaak. Daar is twee orale velflappe wat die velums genoem word, een aan elke kant van die mond, wat met trilhaartjies bedek is. In sommige spesies kom die pre-orale tentakels op die voorkant van die velums voor, maar is afwesig in ander. Daar is egter tien pare pre-orale tentakels in Neopilina rebainsi.
Die mondopening is ‘n horisontale gleuf wat voor deur ‘n dik, V-vormige voorste lip begrens word en agter deur ‘n heelwat kleiner agterste lip met die vertakte postorale tentakels. Die mond is toegerus met vyf pare spiere weerskante van die mond.
Voet
Die ronde voet (wat nie in die skets aangedui word nie) kom soos dié van koppotiges en kewerslakke, onder die liggaam voor. Alle ekstante monoplakofore het agt paar voetspiere wat die liggaam onder aan die skulp vasheg (hierdie spiere word in baie betroubare bronne die pedal rectractor muscles, shell retractor muscles, maar ook lateral en medial pedal muscles genoem, en dan sal sommige soos Ponder et al., (2020) daarna ook as shell muscles verwys. Ek het besluit om die eenvoudigste en mees beskrywende naam, naamlik voetspier, hier te gebruik al heg hierdie spiere aan die binnekant van die skulp vas). Die voetspiere verbind die voet met die skulp en is rondom die interne organe gerangskik. Elke spier verdeel in twee spierbundels van sy vashegtingspunt op die skulp – die mediale en laterale voetspiere. Die mediale voetspier strek bo-oor die onderste senuweekoord en vertak verder in die middel van die voet, terwyl die laterale voetspier langs die onderste senuweekoord verbystrek en in die rand van die voet verder vertak. ‘n Stel kringspiere omring die voet en die mantelgroef, maar is nie aan die skulp geheg nie. Drie stelle spiere – die preorale spier en die voorste en agterste orale spiere, verbind die kop met die skulp.
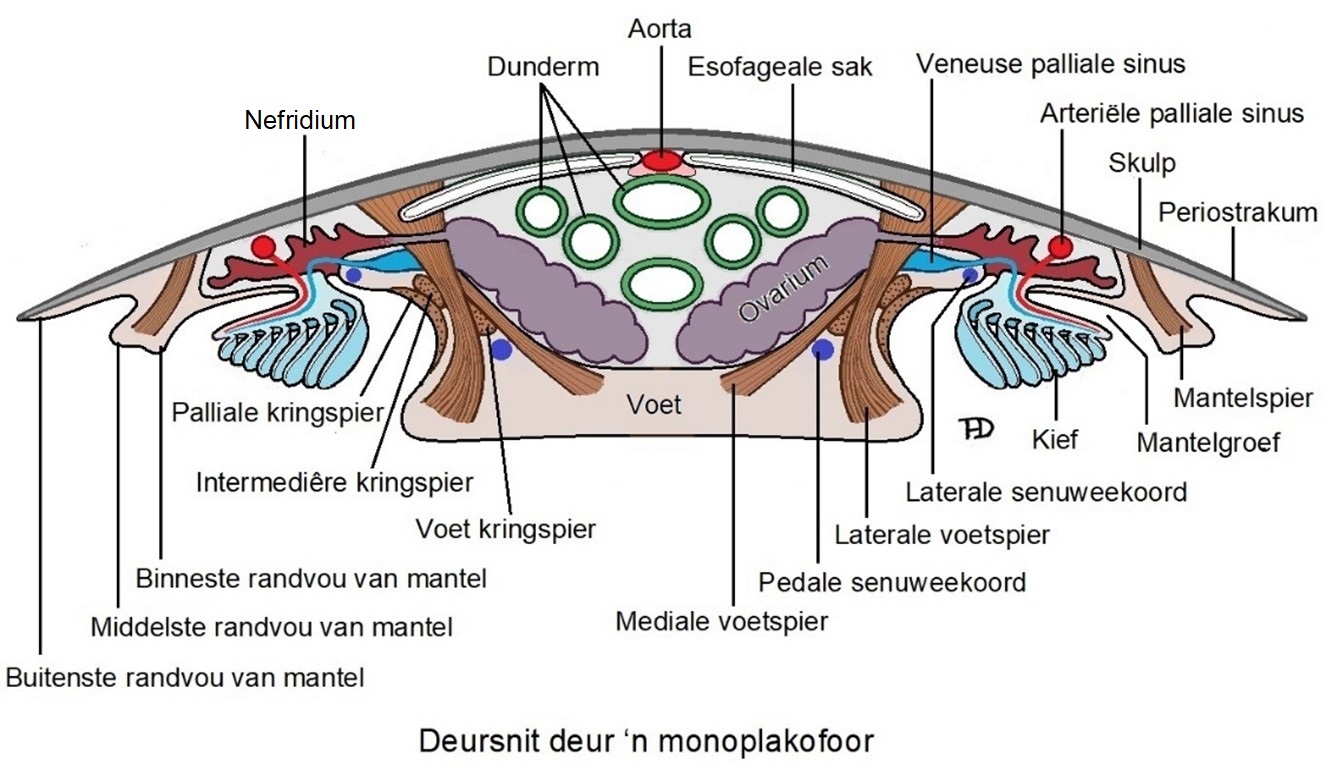
Mantelgroef en kiewe
Monoplakofore het ‘n wye mantelgroef wat tussen die sywand van die voet en die onderste mantelrand voorkom. In meeste monoplakofoorspesies kom daar vyf tot ses paar kiewe in die mantelgroef voor, terwyl ander weekdiere meestal net een paar kiewe besit. Die piepklein Micropilina arntzi het egter net drie paar kiewe. Die kiewe van monoplakofore is ietwat gespierd en beweeg sodat water verby die kiewe kan spoel. Die kiewe het egter ‘n dik epiteel, klein bloedruimtes en digte bedekking van trilhaartjies wat daarop dui dat dit nie so effektief as respiratoriese organe as die kiewe van ander weekdiere funksioneer nie en wat sal verklaar hoekom daar soveel kiewe in monoplakofore voorkom. Die kiewe ventileer egter ook die oppervlakte van die breë mantelgroef bo dit wat as bykomende respiratoriese oppervlakte optree. Daar is ook ‘n noue assosiasie tussen die kiewe en die nefridiums. Die nefridiums speel ook ‘n rol in gaswisseling en word met groter bloedvate aan die aurikels verbind as die bloedvate wat die kiewe met die aurikels verbind.
Spysverteringstelsel
Die mond open onder die kop. Agter die voorste lip is die enkele dik kaak geleë wat weerskante van die mond verbysteek. Die agterste lip (nie op die skets uitgebeeld nie) is agter die mond geleë. Soos in meeste ander weekdiere kom daar ‘n radula binne die mondholte voor. Klein buislose speekselkliere open in die voorste deel van die mondholte.
Die radula, kaakspiere, kraakbene en ondersteuningstrukture rondom die mond van monoplakofore is baie soortgelyk aan dié van die kewerslakke. Die radula van monoplakofore besit rye van 11 tande teenoor die 17 tande van kewerslakke.
Die slukderm bestaan uit ‘n voorste, middelste en agterste deel. Die voorste deel word deur ‘n dorsale gesilieërde voedselkanaal en ‘n paar lengtelopende voue weerskante gekenmerk. Die middelste deel is weerskante aan twee paar groot esofageale sakke met mikrovilli gekoppel. Die agterste deel is ‘n eenvoudige buis wat in die maag open.
Die kleinerige maag is driehoekig in vorm en is in die middel van die liggaam geleë. Die twee lobbe van die ventraal-geleë spysverteringsklier is met ‘n enkele groot opening met die voorste deel van die maag in verbinding. Die spysverteringsklier is, soos in ander weekdiere, met twee tipes selle uitgevoer, maar anders as in ander weekdiere kom daar groot kosdeeltjies in die lumen van die spysverteringsklier voor. Daar kom geen maagskild, sorteerareas of tiflosole in die maag van monoplakofore voor nie.
‘n Sekum is in die agterste deel van die liggaam geleë en is in meeste monoplakofore kort. Die lumen van die sekum bevat epiteelselle met lang mikrovilli.
Die dermkanaal van monoplakofore, wat ek nie in die skets uitbeeld nie (sodat mens die segmentaal-gerangskikte organe beter kan sien), is relatief lank en maak vier tot ses antikloksgewyse lusse voordat dit by die agtertoe-gerigte anus uitkom. Die dermkanaal is rond in deursneë en die binneste plooilose oppervlakte is bedek met gesilieërde epiteel. Die rektum, wat nie struktureel van die dermkanaal onderskei kan word nie, gaan deur die perikardium en eindig by die anus. Nie-voedende monoplakofore ledig hul dermkanaal binne 24 uur.
Renoperikardialestelsel
Die grootste deel van die liggaamsholte van monoplakofore vorm ‘n hemoseel, terwyl die selomiese ruimtes, wat met mesodermale endoteel uitgevoer is, in die perikardium, nefridiums en gonades voorkom. Die liggaamsholte bevat verskeie tipes hemosiete met porieselle (rhogosiete) wat die mees algemene tipe is, terwyl bloedselle, amebosiete en fibroblaste ook voorkom. Die fibroblaste vorm die kollageenvesels wat ‘n groot deel van die liggaamstruktuur vorm.
Die hart is in die agterste, boonste (posterodorsale) deel van die monoplakofoor se liggaam geleë – buiten vir die piepklein Micropilina wat nie ‘n hart besit nie. Die rektum beweeg deur die perikardium en ventrikel wat dan die verkeerde indruk skep dat dit die hart in twee verdeel. Die aorta strek van die ventrikel agter en eindig voor naby die voorste deel van die spysverteringstelsel. Twee pare aurikels van die hart ontvang bloed van die agterste drie paar nefridiums.
Daar is tussen drie tot sewe paar nefridiums in monoplakofore – die groter spesies het meer en die kleineres het minder. Die nefridiums verskil in vorm, is nie aanmekaar gekoppel nie en elkeen open langs die basis van ‘n kief. Net vir herinnering – die hooffunksie van die kiewe van monoplakofore blyk nie om van ‘n respiratoriese aard te wees nie, maar dien eerder as ventilleerders – soos waaiers wat water verby die respiratoriese oppervlaktes bo dit te laat spoel, maar tergelykertyd dan die metaboliese afvalstowwe wat die nefridiums afskei, in die stroom wat geskep word, weg te spoel.
Voortplanting
Die geslagte is in meeste monoplakofoorspesies geskei en individue het twee paar ovaria of testes wat met geslagsbuisies of gonodukte wat binne die lumen van die die derde en vierde paar nefridiums open (in die skets noem ek die geslagsorgane bloot gonades want dit is die versamelnaam vir ovaria of testes). Die piepklein Micropilina arntzi het egter ‘n enkele gonade wat beide eiers en sperms vorm en wat hermafrodities is. Die sperms of eiers word saam met die urine deur die nefridiumsbuisies na buite uitgeskei waar uitwendige bevrugting plaasvind. Monoplakofore se gonades is dorsaal in die liggaam geleë eerder as ventraal soos in die geval van die aplakofore en kewerslakke.
Embrios vorm in Micropilina arntzi, terwyl ander monoplakofore sperms en eiers na buite vrystel en waar uitwendige bevrugting plaasvind. Die grootte van die eiers dui daarop dat dit met dooier toegerus is om die ontwikkeling van die embrio buite die wyfie te onderhou. Die sperm-morfologie is tipies van diere wat eiers in water – ektakwaties (in die waterkolom) of entakwaties (in die mantelgroef van die wyfie) – bevrug. Die groot volume van sperms in manlike individue dui egter op ektakwatiese bevrugting.
Senuweestelsel en sintuie
Monoplakofore se sentrale senuweestelsel bestaan uit ganglionsa en senuwees. Die ganglions is klein en die hoofsenuwees is koordvorming. ‘n Serebrale kommisuur verbind die serebrale ganglionsa. Senuwees wat uit elke serebrale ganglion ontspring innerveer die voorste mantelrand rondom die kop en die mondstreek.
Die pedale ganglions, wat met mekaar deur die postorale kommisuur verbind is, is ietwat na die middel en onder die serebrale ganglions geleë. Die twee klein bukkale ganglions is voor en bo die serebrale kommisuur langs die mond geleë.
Twee paar ventrale hoofsenuweekoorde boog rondom die liggaam agtertoe, die laterale senuweekoorde strek buite-om die voetspiere en die voetsenuweekoorde loop deur die voet aan die binnekant van die voetspiere. Die senuwees wat die transversale verbindings tussen hierdie twee senuweekoorde vorm, strek tussen die voetspiere deur. Die ventrale senuweekoorde vorm ‘n dubbel sirkel en is agter die voet aanmekaar verbind, terwyl dit voor deur middel van die serebrale en voetganglionsa aan die sirkumesofageale ring gekoppel is. Die ringvormige onderste senuweekoorde van monoplakofore verskil van dié van die leervormige senukoorde van kewerslakke en lyk meer soos dié van patellogastropode.
Die pre-orale tentakels het waarskynlik ‘n chemosensoriese funksie. Sekere navorsers spekuleer dat dit moontlik ook fotosensitief kan wees maar, weens die dieptes waarop hierdie diere in die see voorkom is dit hoogs onwaarskynlik, wat ook natuurlik die rede is hoekom hulle nie oë het nie. Dit is nog onbekend wat die funksie van die kompleks vertakte postorale tentakels is, maar ek dink dat dit moontlik ‘n rol in die waarneming van feromone kan hê, want daar was eerstens nog nie ‘n sintuig ontdek in die monoplakofore wat daarvoor verantwoordelik is nie en aangesien hierdie diere in totale duister leef sal dit belangrik wees om ‘n sintuig te hê waarmee hulle paringsmaats kan opspoor. Monoplakofore se subradula orgaan is groot en vervul ‘n chemosensoriese funksie om voedsel en die substraat se chemiese samestelling waar te neem. Osfradieë, die chemiese sensore wat in meeste weekdiere die waterkwaliteit by die kiewe monitor, is ook afwesig in monoplakofore. Anders as die kewerslakke, het monoplakofore ‘n paar statosiste wat verantwoordelik vir die waarneming van aantrekkingskrag is.
Ekologiese rol en gedrag
Ekstante monoplakofore kom wêreldwyd op die seevloer op dieptes van 174 m tot 6 500 m voor. Sommige kom op harde oppervlaktes, insluitende mangaan en fosfaatnodules wat op die seevloer vorm voorkom, terwyl ander in sand en sagte modder voorkom. Dit blyk dat die vroegste Palasosoïese monoplakofore in vlakker water van minder as 100 m diep voorgekom het, maar dat hulle vanaf die Siluur (na dieper water verhuis het (Lindberg, 2009), waarskynlik weens kompetisie met buikpotige weekdiere. Sewe monoplakofoorspesies kom in die diepste dele van die see (4 000 – 6 000 m) voor. Monoplakofore word al hoe groter soos die diepte van die see toeneem en die grootste monoplakofoorspesies kom ook in die diepste dele van die see voor (Lindberg, 2009).
Monoplakofore se maag, spysverteringsklier en dermkanaal bevat meestal die oorblyfsels van mikroskopiese foraminifere, diatome en dinoflagellate. In sekere spesies kom daar egter ook oorblyfsels van mikroskopiese radiolarieë, foraminifere, spons spikula, rondewurms, borselwurm-borseltjies, minerale en organiese materiaal in die spysverterigstelsel voor, wat op substraatvoeding dui.
Bakteriële simbionte kom in die dak van die mantelgroef, in en om die kop, die kiewe, die sywand van die voet en op die postorale tentakels voor. Tot dusvêr weet ons nie wat die funksie van hierdie bakterieë is nie.
Filogenetiese posisie
Molekulêre studies (verwys na analises van DNS, proteïene en ander organiese molekules) dui aan dat ekstante monoplakofore en kewerslakke naaste aan mekaar verwant is (Wilson et al., 2010). Die naam Serialia is voorgestel vir die groep waaronder die monoplakofore en kewerslakke val (Giribet et al., 2006). Dit blyk waarskynlik te wees dat van die oudste weekdiere monoplakofooragtig was en dat die kewerslakke vanuit ‘n monoplakofoorvoorouer ontstaan het (Lindberg, 2009).
Daar is nie konsensus onder paleontoloë en malakoloë ten opsigte van die taksonomiese posisie van sekere Paleosoïese fossiele wat as monoplakofore geïdentifiseer is nie. Terwyl daar konsensus is dat die Paleosoïese familie Tryblidiidae wel aan die monoplakofore verwant is. Die taksonomiese posisie van ander Paleosoïese taksa is onduidelik. Waarteenoor monoplakofore redelik algemeen in die fossielrekord van die Kambrium Periode voorkom, word hulle al skaarser in die Ordovisum en Siluur Periodes en is hulle afwesig in die fossielrekord van die Mesosoïkum Era, terwyl slegs een fossielspesie uit die Kainosoïkum Era bekend is. Hierdie gaping in die fossielrekord het eerder te doen met die bewaring van die sedimente in die afsettingsomgewings waarin hulle voorgekom het, toe hulle van vlakwater na diepwater verskuif het, as wat dit ‘n aanduiding van hul volopheid toentertyd is.